
Page 29 - Fall 2011
P. 29
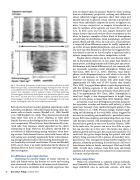
Fig. 4. Narrowband spectrograms of high-arousal, screams produced by an adult female macaque (top), and Arnold Schwarzenegger shouting the words “shut up,” as recorded from the movie Kindergarten Cop (1990). The macaque screams deter- ministic chaos throughout, with no apparent evidence of formant patterning. Schwarzenegger’s shout initially shows regular vocal-fold vibration (the “uh” sound from “shut”), but then gives way to unstable, irregular, and likely chaotic action. Schwarzenegger’s initially distinctive voice quality is readily apparent in the stable portion, but disappears when the source energy becomes unstable. Both species have been tested in playback experiments under naturalistic conditions, with adult females hearing either har- monic calls or chaotic screams (Rendall et al., 1996; Rendall et al., 1998; Rendall et al., 2009). These listeners heard sounds from either their own or others’ offspring, or from adult females that were either biological kin or non-kin. Outcomes were unequivocal. When listeners heard harmonically struc- tured calls, their responses clearly depended on the caller’s relationship to them. However, the subjects showed little or no evidence of differentiating among vocalizers when hear- ing screams. Naïve human listeners tested with rhesus calls in a lab setting were similarly significantly better at discriminat- ing among individual callers when hearing harmonically ver- sus chaotically structured vocalizations (Owren and Rendall, 2003). As yet, there is no ready explanation for the absence of filtering effects in these screams, a puzzle that begs for fur- ther investigation. Sex differences and sexual selection Examining the possible impact of sexual selection on male and female voices has become an active and exciting area of research. Sex differences in human vocal characteris- tics are, of course, so familiar from everyday experience that they are almost taken for granted. However, when working from an evolutionary perspective, noticing such differences almost reflexively triggers questions about their origin and possible function. In general, sexual selection is proposed to occur when individuals compete for access to opposite-sex mates (intrasex competition), or compete to be selected as a mate by members of the opposite sex (mate-choice competi- tion). In both cases, one sex may acquire distinctive and unique features that need not have direct counterparts in the other. In humans, examples of these kinds of dimorphisms include body-fat distribution, facial morphology, and beard growth (Boyd and Silk, 2011). In such cases, sexual selection is suspected when differences cannot be readily explained as an artifact of more global dimorphisms, such as in body-size. The next step then becomes to show that the exaggerated fea- tures found in one sex or the other play a significant role in intrasex competition, mate-choice competition, or both. Possible effects of body-size on the voice become impor- tant in that primate males are, in fact, larger than females in many species, including humans and all four great ape species. Furthermore, male-female differences in F0 are common with- out necessarily exceeding overall dimorphism (Mitani and Gros-Louis, 1995; Ey et al., 2007). However, vocal dimor- phisms can be disproportionate as well, which is the case for both F0 and formants in baboons (Rendall et al., 2004). Outcomes for humans are similar, with adult males being approximately 8% taller and 15-20% heavier than females (Puts, 2010). Laryngeal dimorphism is quite disproportionate, with the vibrating segments of the adult vocal folds being about 60% longer in males than in females, which lower speak- ing F0 by approximately 50% (Titze, 1994). Dimorphism in vocal tract length is also disproportionate to height, being about 15–20% greater in males (Fant, 1960; Goldstein, 1980). In humans, vocal-tract development proceeds along sim- ilar trajectories in males and females until puberty, at which point boys famously show marked laryngeal growth (Titze, 1994; Harries et al., 1998). Physical changes include length- ening and thickening of the vocal folds, effects triggered by increases in circulating sex steroid levels—particularly testos- terone. Both masculinizing and feminizing effects are classi- cally hormone-related, with dimorphism resulting from dif- ferential tissue growth in one sex or the other (Dixson, 2009). In the male voice, the process can occur in as little as a year, but can also take up to five years. The larynx also shows a pubertal growth spurt in girls, but much more modestly. The vocal-tract also grows longer during this period, with male puberty being associated with a process of secondary laryn- geal descent. This laryngeal lowering thereby lengthens the pharynx, ultimately positioning the male larynx a full verte- bra’s distance below its female counterpart (Fitch and Giedd, 1999). Overall, then, evidence from both male and female anatomy and vocal acoustics are indicative of sexual selection in human vocal production. Intrasex competition Within-sex competition is common among mammals, most frequently between males (Puts, 2010). As a rule, the larger individual wins in male-male contests, with many 28 Acoustics Today, October 2011
