
Page 70 - Winter 2020
P. 70
FROG EARS
the shoulder girdle, to the opercularis muscle, to the operculum, and finally to the inner ear. Action of the operculum and opercularis muscle may also protect the ear from damage during respiration and vocalization by shunting vibrations away from the inner ear. Vibrations of the operculum and the footplate are coupled, but how this coupling affects the hypothesized functions of the operculum is not well understood (Mason, 2007).
Frogs can also detect sounds using their lungs in a manner that is analogous to the use of the swim bladder in bony fishes (Popper and Hawkins, 2018). Narins et al. (1988) demonstrated that an area of the lateral body wall overlying the lungs of the Puerto Rican coqui frog Eleutherodactylus coqui vibrates in response to airborne sound. When sound waves hit this region, the resulting vibrations are funneled through the lungs and glottis to the mouth cavity, Eustachian tubes, and middle ear cavity to the oval window. Many earless species rely on the lung pathway to detect sounds (Mason, 2007).
Bone conduction is also an important transmission pathway. Gardiner’s Seychelles frog (Sechellophryne gardineri) is a small animal lacking a middle ear cavity and a columella. It has poorly developed lungs and an ossified operculum; still, it responds to sounds and it produces advertisement calls (Boistel et al., 2013). The mouth cavity of this frog resonates at around 5,700 Hz, a spectral peak in the male’s advertisement call. This resonance is transmitted to the inner ear through bone conduction (Boistel et al., 2013).
The Unique Frog Inner Ear
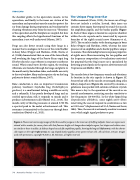
Unlike mammals (Puria, 2020), the inner ear of frogs does not include a cochlea. Instead, their inner ear contains three organs that respond to sound: the saccule, the amphibian papilla, and the basilar papilla (Figure 4). Each of these organs is located in a separate chamber within the otic capsule and is innervated by separate branches of the eighth cranial nerve (Simmons et al., 2007). The saccule is similar to the hearing organ in bony fishes (Popper and Hawkins, 2018), whereas the joint presence of an amphibian and a basilar papilla is unique to anurans. The relationship between response properties of eighth nerve fibers innervating the two papillae and the frequency composition of vocalizations motivated the proposal that the frog’s inner ear is specialized for detecting spectral peaks in the species advertisement call (Capranica and Moffat, 1983).
The saccule detects low-frequency sounds and vibrations. Its location in the otic capsule is shown in Figure 1C. Sensory hair cells in the saccule are arranged along a flat, kidney-shaped area (Figure 4A) covered by otoconia, a gelatinous mass packed with calcium carbonate crystals. This mass is key to the operation of the saccule as an inertial accelerometer, restricting saccular sensitivity to low frequencies (20-300 Hz). In the white-lipped frog (Leptodactylus albilabris), some eighth nerve fibers innervating the saccule respond to accelerations as low as 0.001 cm/s2 (displacements of 10 Å; Narins and Lewis, 1984). Thus, the saccule is extremely sensitive to seismic cues, which might signal predators or prey.
Figure 4. Fluorescent microscopy images of the three auditory organs in the inner ear of bullfrog tadpoles. Inner ear organs were stained with a marker for sensory hair cells that fluoresce bright red. Images show tadpole organs, but shapes and relative sizes are similar to those in adults. A: kidney-shaped saccule; B: amphibian papilla, showing the long tail (left arrow), with the shorter club region on the right (right arrow); C: cup-shaped basilar papilla; arrow points to hair cells. All scale bars, 100 μm. Images used with permission, copyright © 2020 A. M. Simmons, all rights reserved.
70 Acoustics Today • Winter 2020
